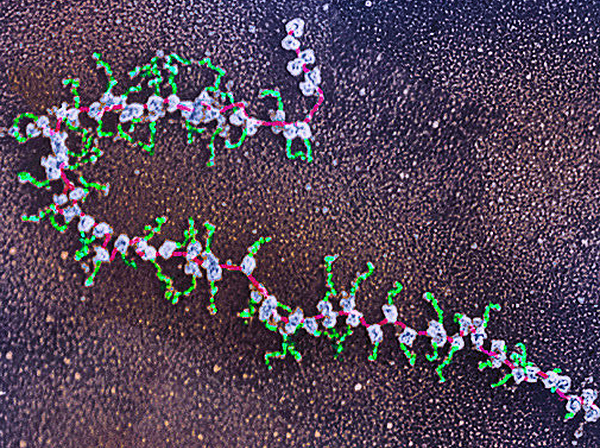 Фото: Dr Elena Kiseleva
Фото: Dr Elena Kiseleva Чем больше конкретных данных нам предоставляет молекулярная биология, биохимия и смежные дисциплины, тем труднее их не только трактовать в заидеологизированных рамках дарвинизма, но и вообще осмыслять в натуралистической парадигме, то есть без привлечения аппарата теории принятия решений в контексте целеполагания и планирования[1][2].
В качестве конкретного примера мне хотелось бы рассмотреть некоторые особенности системы размножения живых существ. Как известно, в процессе размножения система репликации производит копирование ДНК, несущую инструкции для синтеза белков, а система трансляции осуществляет обработку этих инструкций. Если не вся, то значительная часть ДНК, непосредственно не кодирующая белок, имеет регуляторные функции[3]. Здесь всё, как и в искусственных распределённых вычислительных системах: кроме самого кода, получателю (в данном случае новообразованному организму) необходимо сообщить, что и как с этим кодом исполняющей программе нужно делать[4].
Вот эта обработка и представляет для нас интерес. Мы не будем касаться таких технических сложностей, как сплайсинг, в результате которого системой обработки из матричной РНК удаляются не кодирующие белок участки (интроны), а белок-кодирующие (экзоны) «склеиваются». Мы затронем только самое основное, не вдаваясь в детали, но лишь вкратце рассмотрев всего два аспекта организации системы размножения: транскрипцию и трансляцию генетического кода, их функции, смысловое различие и конкретную реализацию.
В процессе обсуждения, я надеюсь, мы увидим, что рассматривать эти функции живых систем вне контекста целеполагания с позиций современных научных данных уже невозможно[5]. В этом смысле мой основной тезис можно сформулировать, перефразировав известные слова биолога Ф. Добржанского: в биологии всё обретает смысл только в свете дизайна.
Терминология
Для начала в целях удобства приведем небольшой список терминов[6], важных для понимания изложенного в этой статье.
• Азотистые основания – гетероциклические органические соединения, производные пиримидина и пурина, входящие в состав нуклеиновых кислот. Для сокращенного обозначения пользуются большими латинскими буквами. К азотистым основаниям относят аденин (A), гуанин (G), цитозин (C), которые входят в состав как ДНК, так и РНК. Тимин (T) входит в состав только ДНК, а урацил (U) встречается только в РНК.
• Аминокислоты – особые органические соединения. Нас интересуют только те из них, которые заняты в процессе синтеза белка. В состав синтезируемых белковых макромолекул включаются 20 стандартных протеиногенных аминокислот, кодируемых генетическим кодом. В некоторых белках присутствуют также нестандартные аминокислоты, получающиеся из стандартных путем посттрансляционных модификаций.
• Белки (протеины, полипептиды) – высокомолекулярные органические вещества, состоящие из так называемых протеиногенных аминокислот, соединенных в цепочку пептидной связью. Аминокислотный состав белков определяется генетическим кодом. Свойства белков определяются последовательностью аминокислотных остатков в полипептидной цепи.
• Генетический код – свойственный всем живым организмам способ кодирования последовательности аминокислотных остатков в составе белков при помощи азотистых оснований (нуклеотидов) нуклеиновой кислоты.
• ДНК – макромолекула дезоксирибонуклеиновой кислоты, несущая информацию, необходимую для воспроизведения живого организма.
• Интеллект – способность принимать решения по достижению цели[7].
• иРНК – информационная РНК, макромолекула рибонуклеиновой кислоты, несущая информацию, необходимую для воспроизведения живого организма. Синтезируется в процессе транскрипции по ДНК.
• мРНК – матричная РНК, см. иРНК.
• Нуклеиновые кислоты – высокомолекулярные органические соединения, биополимеры, образованные остатками нуклеотидов. Нуклеиновые кислоты ДНК и РНК присутствуют в клетках всех живых организмов[8] и выполняют важнейшие функции по хранению, передаче и реализации наследственной информации.
• Нуклеотиды – см. Азотистые основания.
• тРНК – транспортные РНК, макромолекулы, необходимые для транспортировки аминокислот к рибосомам в процессе синтеза белка.
• Принятие решений – выбор из альтернатив[9].
• Рибосома – макромолекулярный комплекс, служащий для синтеза белка по заданной матрице на основе генетической информации, предоставляемой мРНК.
Обработка генетического кода в живых организмах
Обработка генетического кода включает две существенно различные фазы: транскрипцию и трансляцию. В соответствии с так называемой «центральной догмой» молекулярной биологии, транскрипция отвечает за первый этап цепочки: ДНК → РНК → белок, трансляция – за второй (рис. 1).
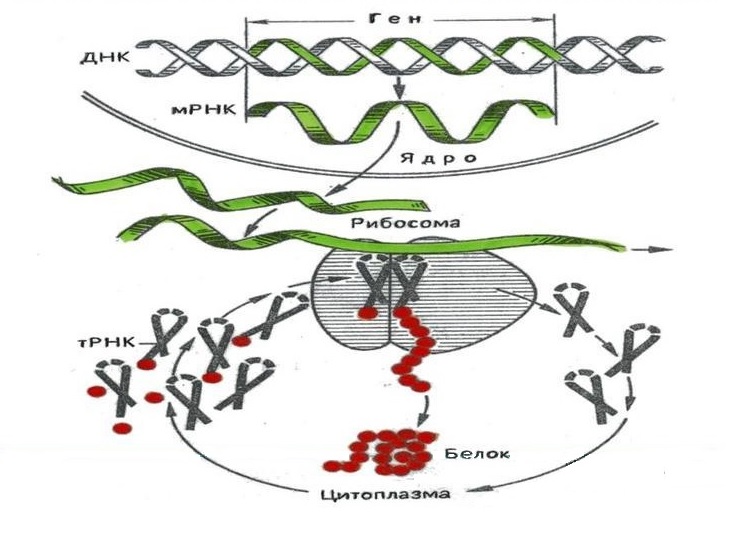 Рис. 1. Общая схема биологического синтеза белка. Источник: МуShared.Ru
Рис. 1. Общая схема биологического синтеза белка. Источник: МуShared.Ru Транскрипция генетического кода
Транскрипция[10] производится в основном с целью синтеза матричной (информационной) РНК, по которой впоследствии в процессе трансляции синтезируется белок. Дополнительными функциями транскрипции является синтез:
• рибосомальных РНК, входящих в состав рибосом;
• транспортных РНК, являющихся основными элементами, осуществляющими при синтезе белка перекодирование информации, заключенной в информационной РНК, с языка нуклеотидов на язык аминокислот;
• РНК, участвующих в репликации ДНК.
Транскрипционная фаза необходима эукариотам, в частности, по той причине, что молекулы ДНК не могут покинуть клеточное ядро, а синтез белка на рибосомах по информационной РНК имеет место в цитоплазме вне клеточного ядра.
Важной функцией транскрипции является регуляторная функция, предоставляющая организму дополнительные возможности по управлению сложнейшими процессами, которые имеют место при размножении на стадии транскрипции. Некоторые РНК являются каталитически активными, что позволяет считать их транскрипцию синтезом конечного продукта в противоположность белкам, синтез которых включает стадию трансляции РНК.
Трансляция генетического кода
Трансляция информации состоит в чтении обработчиком некоторой материальной конфигурации как последовательности знаков (символов) и интерпретации ее по правилам, заданным в определенном информационном контексте. Интерпретация заключается в том или ином физическом эффекте, который производится обработчиком (например, выдача сообщения на экран, если речь идет о выполнении компьютерной программы, или образование нового химического соединения, как в процессе синтеза белка). Этот эффект по отношению к знаку называют референтом или денотатом (букв. «тем, что обозначается» знаком). Чрезвычайно важно то, что смысловое содержание интерпретации определяется не самим знаком, а соответствующим протоколом. Знак лишь инициирует собственную обработку. Таким образом, между знаком и референтом в системе трансляции существует только логическая связь. Эта связь осуществляется через протокол. Физическая реализация протокола может быть различной[11].
Трансляция[12] информационной РНК состоит в синтезе белков, который осуществляется на рибосомах интерпретацией триплетов азотистых оснований (нуклеотидов) информационной РНК с помощью особых транспортных РНК (тРНК). Интерпретация производится по правилам, называемым генетическим кодом. Эти правила устанавливают логические соответствия между тройками нуклеотидов информационной РНК и аминокислотными остатками, которые будут вставлены в синтезируемый линейный полипептид, впоследствии укладывающийся в трехмерную молекулу белка. Трансляция предполагает транспортировку нужных по генетическому коду аминокислот с помощью транспортных РНК к рибосомам, отгрузку аминокислот и их присоединение к полипептиду.
Знаками здесь выступают последовательности кодонов информационной РНК, правилами – генетический код, референтом – полипептид. Информационным контекстом является создание всего множества белков (т.н. протеома) дочернего организма. Что касается степени сложности белкового синтеза, то для иллюстрации достаточно привести одну цифру: размер протеома человека составляет порядка 20 тысяч различных видов белка[13]. Скорость синтеза белка в живых организмах – регулируемая величина с максимальным значением порядка десятков аминокислотных остатков в секунду[14].
Процессы записи/считывания и интерпретации между собой физико-химически не связаны
Удивительная особенность трансляции информации вообще (и в живых системах в частности) состоит в том, что процессы записи/считывания и интерпретации между собой физико-химически не связаны. Напротив, одним из условий успешной трансляции генетического кода именно как кода является как раз независимость физико-химического представления (в форме молекулы информационной РНК) инструкций по построению белка от их интерпретации (в форме полипептида)![15] Аналогичным образом и представление понятий в виде записи слов на определенном языке абсолютно безразлично относительно законов природы и не определяется ими. Поэтому существование самой системы трансляции невозможно объяснить натуралистически, то есть исключительно в терминах движения частиц вещества системы. Действительно, натурализм, оперируя лишь случайностью и закономерностью, априори исключает категорию принятия решений со стороны агента, а без привлечения аппарата теории принятия решений происхождение живых организмов, являющихся не чем иным, как самовоспроизводящимися системами принятия решений[16], объяснить принципиально невозможно! Современный американский философ-атеист Алекс Розенберг неправ, когда утверждает[17], что в мире ни один сгусток материи не обозначает никакой иной сгусток материи. В системах трансляции именно это имеет место: одни конфигурации материи (знаки) обозначают другие (референты). Биологические системы, в которых трансляция кода – объективно установленный научный факт, – ярчайшее тому подтверждение[18][19].
Для того, чтобы лучше уяснить, что имеется в виду, рассмотрим простой пример. Водитель машины, движущейся по городу, видит дорожный знак «Проезд закрыт» и соответствующим образом регулирует движение транспортного средства (скажем, поворачивает вправо). Движения мышц водителя не зависят от расположения частиц краски, нанесенных на металлическую основу знака, в том же смысле, в каком движение отпущенного с высоты шарика зависит от наличия гравитации. Дорожный знак налагает граничные условия на движение мышц водителя, что предполагает интерпретацию водителем смысла знака в соответствии с правилами дорожного движения. В случае же шарика никакой интерпретации нет, есть лишь его механическое движение в поле тяготения. Водитель волен нарушить требование знака, в то время как шарик никак не может «воспротивиться» притяжению Земли! Заметим, наконец, что возможность создания семантически нагруженных конфигураций материи (знаков) возникает только при условии множественности безразличных положений равновесия в системе. Тот же водитель, оказавшись на месте шарика, так же, как и шарик, будет двигаться вниз под действием силы тяжести, хочет он этого или нет.
Система трансляции генетических инструкций, как и дорожный знак, лишь налагает граничные условия на движение вещества в системе. Отличие ее от дорожного знака заключается лишь в особенностях реализации: граничными условиями на движение частиц системы (иРНК, рибосомы, полипептид) являются правила трансляции, физически реализованные с помощью особых ферментов (арсаз), нагружающих нужными аминокислотами макромолекулы тРНК для синтеза полипептида. То, какой аминокислотой будет нагружена данная тРНК, никак не зависит от последовательности нуклеотидов в информационной РНК. Нужная по генетическому коду тРНК разгрузится на рибосоме с последующим присоединением транспортируемой ею аминокислоты к полипептиду только потому, что тРНК, как вилка в розетку, войдет своим антикодоном в кодон иРНК. «Не те» тРНК в розетку не воткнутся! Это адаптер в чистом виде (рис. 2).
Также важно осознавать, что для обеспечения репликации организма необходимо воспроизвести и саму систему трансляции. Поэтому инструкции по воспроизведению системы трансляции закодированы тем же кодом, что используется самой этой системой для воспроизведения остального организма! Попытки натуралистически[20] объяснить данное наблюдение – не что иное, как хождение по порочному кругу.
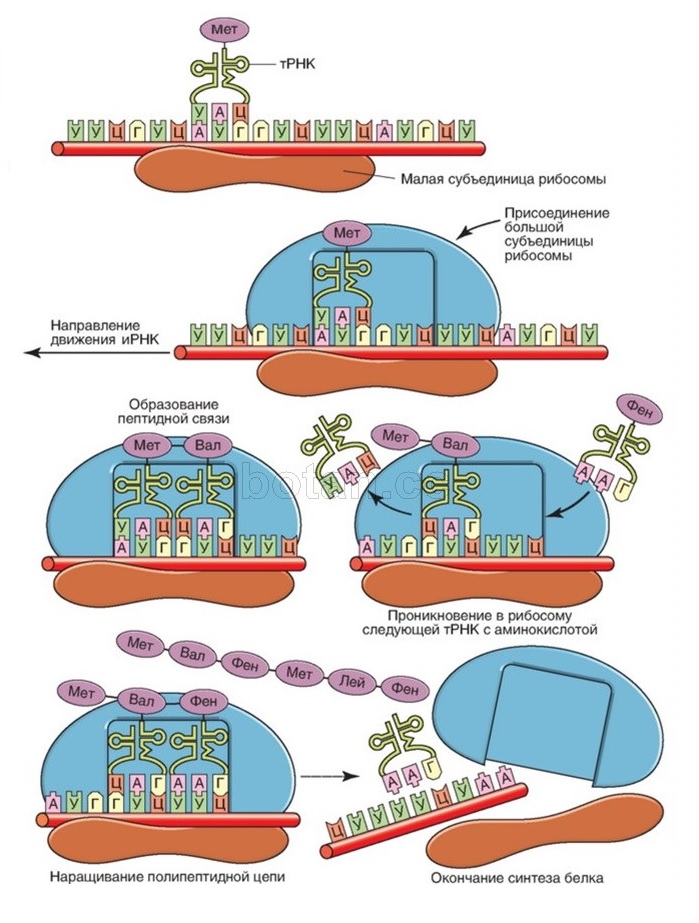
Как сказал один проницательный интернет-комментатор при обсуждении знаменитой адаптерной гипотезы[21] Френсиса Крика: случайность не строит адаптеров! Он прав: для организации движения материи (в частности, в форме систем трансляции информации) необходимо предвидение, целеполагание, планирование и способность принятия решений, иными словами – интеллект! И это не голословное утверждение верующих ученых. Напротив, это вывод[22], проистекающий из обобщения наблюдений: примеры трансляции информации в природе исчерпываются лингвистическими и биологическими системами. Способность интеллекта создавать лингвистические системы – естественные языки и языки программирования, математика, шифры с их ключами и пр. – общеизвестна[23]. С другой стороны, в неживой природе трансляции информации не наблюдается[24]. И этого следовало ожидать, поскольку неживая природа лишена интеллекта: она не принимает решений, не ставит целей, не планирует и не выбирает из альтернатив.
Для создания системы трансляции генетической информации нужен был интеллект
Таким образом, совершенно естественно предположить, что и для создания системы трансляции генетической информации также нужен был интеллект! А если это так, то интеллект должен был быть ответственным за организацию всего живого, поскольку без воспроизведения, а значит, и без трансляции генетической информации, общей для всех организмов, жизнь немыслима.
Суть гипотезы дизайна биосистем
Мы подошли к самой сути гипотезы дизайна биосферы. Эта гипотеза по своему смыслу является предположением о начальной функциональной организации живого. По этому предположению, неинтеллектуального физико-химического пути от неживого к живому не существует. Исходя из анализа биологических систем, можно сделать единственно верный вывод: происхождение жизни, насколько бы более простыми ни являлись организмы первого поколения по сравнению с современными, изначально потребовало организации, в частности в виде особым образом реализованных символьных граничных условий с целью обеспечения трансляции кода.
Трансляция информации – не результат эволюции, а ее основная причина
В этом кардинальное отличие и существенное преимущество гипотезы дизайна по сравнению с оторванными от реальности натуралистическими гипотезами постепенного эволюционного формирования системы трансляции только за счет неинтеллектуального химического синтеза. Именно трансляция информации предполагает (при наличии иных важных условий) дальнейшие эволюционные движения живого (безотносительно реальных возможностей биологической эволюции, которые обсуждать сейчас мы не будем[25]), а не наоборот! Организационные системные эффекты являются причиной последующих эволюционных движений биосистем, а не наоборот. В утверждении о том, что трансляция информации представляет собой результат эволюции, а не основную причину последней (каковы бы ни были реальные возможности эволюционирования самовоспроизводящихся открытых систем), содержится категориальная ошибка чудовищного масштаба современных абиогенетических моделей неинтеллектуального синтеза.
Суть аргумента организации состоит в том, что граничные условия и само движение частиц материи, составляющей физическую систему (в данном случае живую), представляют собой две совершенно различные категории. Эпистемологическое разграничение категорий движения материи и граничных условий, накладываемых на него, известно со времен Исаака Ньютона и составляет, по мнению американского биофизика Х. Патти[26], главный вклад гениального английского ученого в развитие науки. Граничные условия никаким образом не зависят от движения частиц системы. Напротив, они задают движение, а не движение частиц системы каким-то непонятным образом организует граничные условия для себя. Сторонники гипотезы неинтеллектуального абиогенетического синтеза ставят телегу впереди лошади. Вспомним хотя бы университетский курс дифференциальных уравнений: из всего (бесконечного) континуума траекторий точечной массы, вытекающего из уравнений ее движения в поле тяготения, именно подстановка граничных условий в уравнения движения выделяет/задаёт одну определенную траекторию, отвечающую реальному движению объекта исследования. Такие категории, как граничные условия и измерение величины[27], отличны от движения частиц материи (от так называемых законов природы) и к нему не сводятся. По этой причине Х. Патти рассуждает о проблеме трансляции и проблеме измерения величин как о проблеме эпистемного сечения: где именно пролегает граница между объектом управления, измерения, трансляции и т.д. и субъектом, осуществляющим эти действия[28].
Таким образом, организация движения частиц материи никаким образом из самого движения не вытекает, как нас постоянно пытаются убедить. Она является внешним независимым фактором по отношению к движению материи. Источник организации движения материи в форме особых символьных граничных условий, что включает чтение/запись и интерпретацию символьной информации, – всегда только внешний по отношению к системе интеллект. Конкретика биофизики и теоретической биологии состоит, в частности, в выявлении этих граничных условий, различных в каждом отдельном случае.
Функциональная дифференциация генетического кода
Еще одной важнейшей особенностью системы воспроизведения живых организмов является дифференцированный подход к генетическому коду. В случае транскрипции генетический код, записанный на ДНК-носителях, воспринимается системой как данные, без интерпретации содержимого. В случае же трансляции фактически тот же код (только в форме иРНК) интерпретируется системой воспроизведения уже как последовательность инструкций по синтезу белка, то есть как программа.
Описанный системный дуализм «программа-данные» в живом, без сомнения, так же как и система трансляции генетического кода, имеет интеллектуальное происхождение, поскольку также относится к категории организации, в отличие от самого движения частиц материи, составляющей систему.
Интересно, что работы Дж. фон Неймана по моделированию живых организмов самовоспроизводящимися клеточными автоматами, исторически предшествовавшие открытию функций ДНК, фактически предсказали этот организационный дуализм живых систем: клеточный автомат в одном случае должен был копировать ленту с описанием структуры автомата как данные (для последующего копирования уже с дочерней клетки), в другом (для воссоздания самой дочерней клетки) должен был интерпретировать содержимое ленты[29].
С интересным обзором результатов математического моделирования биосистем клеточными автоматами можно ознакомиться в статье Koza John R. Spontaneous Emergence of Self-Replicating and Evolutionarily Self-Improving Computer Programs, мой критический анализ которой был недавно опубликован блогом UncommonDescent.com[30].
Обсуждение конкурирующих натуралистических гипотез происхождения жизни
Согласно гипотезе РНК мира, являющейся одной из наиболее популярных на данный момент гипотез происхождения жизни, упомянутая мною выше цепочка от ДНК до белка является молекулярным реликтом, доставшимся нам от гипотетического пребиотического мира РНК[31]. Эта гипотеза постулирует, что как функцию хранения генетической информации, так и катализ химических реакций первоначально выполняли ансамбли молекул рибонуклеиновых кислот. Тем не менее, хранение генетической информации уже предполагает считывание ее и последующую интерпретацию. С точки зрения происхождения самой системы трансляции РНК мира (а равно и родственные ей гипотезы ДНК мира или мира белка), на мой взгляд, ничего не дает. Разница лишь в том, что предполагается, будто трансляция выполнялась с помощью исключительно РНК. Сведение типа организации, наблюдающегося в современных нам живых системах, к системам на основе исключительно РНК, таким образом, лишь на первый взгляд представляется упрощением, поскольку вопрос о том, как возник механизм трансляции кода, остаётся без ответа.
Другой популярной гипотезой является гипотеза добиотических метаболических сетей[32]. Эта гипотеза базируется на том, что в своей основе обменные сети всех организмов включают соединения углерода и структурно схожи между собой, из чего делается гипотетический вывод о раннем эволюционном происхождении этой обменной углеродной основы. Однако гипотеза метаболических сетей в отношении трансляции информации, важном в нашем обсуждении, столь же спекулятивна, сколь и РНК мир, поскольку предполагается, что ключевыми в происхождении жизни являлись физико-химические ограничения. Исключительная роль, фактически роль интеллекта, в рамках этой гипотезы отводится законам природы и особым начальным условиям. Но, как мы уже видели, такому отведению не существует сколь-нибудь серьезного эмпирического обоснования.
В случае РНК-репликаторов неочевиден переход от простого копирования к интерпретации кода. Что касается гипотезы обменных сетей, то непонятно, как работающая сеть реакций может вообще развиться до степени сложности, которой характеризуются метаболические сети современных организмов.
Ни та, ни другая гипотеза не отвечают на главный вопрос о происхождении первого транслятора информации, то есть на вопрос о происхождении эпистемного сечения: измеритель/измеряемое, управляющий контур/управляемая подсистема, движение материи/граничные условия, код/интерпретатор. С другой стороны, единственным достоверно известным из опыта источником эпистемного сечения является внешний по отношению к системе интеллект. Никакие физико-химические ограничения сами по себе не способны обеспечить обособление компонентов системы, выполняющих роли кода и транслятора кода. Повторю, что, на мой взгляд, это так именно в силу самой сути трансляции информации, одним из условий которой является физико-химическая независимость эффекта трансляции от конфигураций материи, выступающих в роли кода.
Как изменяется научная повестка с введением гипотезы дизайна
В заключение мне хотелось бы вкратце коснуться такой интересной проблемы, как конкретные выводы из гипотезы дизайна. Коль скоро речь заходит о научном моделировании, интересны именно следствия из формулируемых гипотез и закладываемых допущений, конкретные с точки зрения моделирования:
• можно ли будет, и если да, то как, экспериментально подтвердить или опровергнуть формулируемые утверждения;
• насколько нетривиальными и интересными будут следствия, вытекающие из них;
• насколько богатыми будут проблемное поле и исследовательская программа, основанные на той или иной гипотезе.
Именно с помощью плодотворных гипотез, открывающих целые направления исследований, наука и движется вперед.
Независимо от того, отдают ли эволюционисты себе в этом отчет, но и они, пусть и неявно, используют допущение о целесообразности в основе функционирования биоты. Всё живое по какой-то причине стремится к выживаемости, включая примеры альтруизма, когда отдельные особи жертвуют собой с целью выживания популяции. Таким образом, при анализе биосистем и эволюционистская программа также предполагает обратную разработку с целью установления рациональных принципов функционирования и строения изучаемых организмов. Так что, на мой взгляд, допущение как минимум об изначальном дизайне не только не испортит картины, но внесет в нее отсутствующую на сегодняшний день ясность и определенность.
Примером подобного подхода может служить вот эта замечательная, на мой взгляд, статья, предлагающая гипотезу универсального генома многоклеточных: Sherman Michael. Universal Genome and the Origin of Metazoa[33]. В отличие от устаревшей и, по большому счету, не оправдавшей себя дарвиновской модели, автор отталкивается от предположения о начальной сложности многоклеточных организмов, допуская, что в кембрии многоклеточные были снабжены общим геномом. При этом не делается никаких предположений об источнике генетической информации, вложенной в организмы кембрийского периода. Сам автор трактует стартовую сложность кембрийских организмов как данность, уподобляя загрузку функциональной информации в биосистемы Большому взрыву, заглянуть «за который» не представляется возможным в рамках данной гипотезы. По Шерману, различия между организмами объясняются различными настройками универсального генома, а именно: выключением и включением соответствующих его участков.
Гипотеза об универсальном геноме приводит к следующим прогнозам.
• У низших многоклеточных доля генетической информации, хотя и присутствующей в их геномах, но не задействованной и поэтому функционально бесполезной, должна оказаться сравнительно высокой; эта доля будет тем меньше, чем выше степень организации того или иного многоклеточного.
• Подобные латентные программы развития возможно будет экспериментальным образом активизировать: например, включить программу развития органов зрения или синтеза антител у морского ежа[34].
Отметим самое важное для нас. Эта дизайн-гипотеза принципиально отлична от дарвиновской модели и приводит к конкретным тестируемым дочерним гипотезам, невозможным в дарвинистской парадигме. Гипотеза универсального генома к тому же объясняет некоторые наблюдения лучше, чем теория Дарвина, предполагающая постепенное прагматически обусловленное развитие от простого к сложному. Например, Шерман лучше объясняет фасеточное зрение у гидр, которое по Дарвину труднообъяснимо (если вообще это возможно), поскольку гидрам оно не нужно, ведь изображение фокусируется вне тела гидры. По Дарвину получается, что бесценные энергетические ресурсы тратились гидрой на зрение практически впустую, поскольку толком она им воспользоваться не может. В случаях, подобных этому, дарвинистам приходится сильно изворачиваться, утяжеляя теорию дополнительными предположениями. По Шерману же всё гораздо проще: в прошлом у гидр могло произойти случайное включение участков универсального генома, ответственных за формирование этого типа зрения, но бывших до того выключенными.
Итак, что касается выработки конкретных рабочих гипотез, как видим, нетривиальных и тестируемых (если не сегодня, то в ближайшем будущем), исследовательская повестка дизайна ничуть не уступает эволюционистской программе исследований. Парадигма дизайна является к тому же более общей по сравнению с эволюционистской, поскольку не исключает эволюцию как таковую, в то время как эволюционизм исключает дизайн априори. Гипотеза дизайна лишь ставит эволюцию на подобающее ей место второстепенного фонового процесса, который хотя и наблюдается, однако не может на практике являться фактором генерации достаточно больших количеств функциональной информации[35] и поэтому не может быть ответственным за всё наблюдаемое биологическое разнообразие.
В рамках гипотезы дизайна, вероятно, возможно восстановить не только внутреннюю логику, лежащую в основе интеллектуальных действий агента в ряде ключевых событий истории биоты (таких, например, как появление многоклеточных, половой дифференциации и пр.), но также временные рамки и последовательность принятия решений по привнесению кардинальной биологической новизны.
Итоги
Подведем итоги нашего обсуждения. Трансляция информации имеет место только в лингвистических и в биологических системах. Способность интеллекта создавать лингвистические системы – достоверный факт. В неживой природе (разумеется, за исключением заведомо искусственных систем обработки информации, созданных человеком) трансляции информации не происходит. Интеллектуальность происхождения биосистем, таким образом, представляется хорошо экспериментально обоснованной гипотезой.
Время работает против «естественников», априори отвергающих гипотезу дизайна биосферы: по мере развития молекулярной биологии и других наук неприятные для сторонников узкопонимаемого натурализма сюрпризы накапливаются всё с большей скоростью. Предлагаемые ими объяснения происхождения жизни, оперирующие лишь случайными и закономерными факторами среды без интеллектуального управления синтезом, всё больше теряют всякую связь с реальностью. Действительно, требуется не только обосновать и детально изложить, каким образом могла сформироваться первая популяция самовоспроизводящихся неоднородных открытых систем с символьной памятью, но и предложить объяснения целого ряда конкретных научных данных. Одним из таких наблюдений, как мы уже видели, является дифференцированный подход живых систем к коду, когда в зависимости от множества факторов практически один и тот же код интерпретируется организмом и как данные, и как программа. Учёт внешнего интеллектуального фактора, напротив, позволяет не только корректно объяснять наблюдения, но и формирует новое нетривиальное проблемное поле для дальнейших исследований.
В заключение я хотел бы выразить благодарность моим коллегам и друзьям из Кембриджского и Брянского технического университетов за ценные обсуждения, замечания и предложения по тексту статьи.




Благодарю за положительный отзыв на статью. Конечно, материал не является чем-то абсолютно новым, об этом давно уже говорится. Новизна лично мне видится в том, как идеи семиотики сегодня прилагаются в биологии.
Хотелось бы только немного уточнить, что Бога все-таки рационально познать нельзя. Можно прийти к вере в Бога от созерцания сотворенного Им мира (в том числе и через науку), как учит св. апостол Павел. Тем не менее, познание Бога происходит благодатно, а не рационально-аналитически. Причаствуется только нетварная Божественная благодать, а не Его сущность.
При образовании кристаллов не происходит трансляции информации, необходимой для воспроизведения живого. Кристаллизация не требует трансляции, а значит, и наличия описания системы во внешней памяти, чтения/записи с неё, протокола. Кристаллы однородны и кроме роста, обеспечиваемого простым матричным копированием, не обладают другими свойствами живых организмов (т.е. неоднородных автономных самовоспроизводящихся систем, реагирующих на стимулы и принимающих решения). Живое, с т.зр. биосемиотики, - это символьно управляемая (то есть особым образом организованная) материя. Если подытожить, то жизнь характеризуется целым набором свойств.
Какая Программа управляет этими процессами для всех существующих кристаллов? ДНК - это тоже кристалл. Так что отнести к неживой материи?
Наши учёные как раз таки и задались этим вопросом: каким образом функционирует ДНК? Приходят у выводу, что ДНК не самодостаточно, а существует в некоем общем поле. По-нашему можно сказать, существует волею Творца. На этой странице подробнее и интереснее: http://tainadiveevo.ru/taina-volnovoy-genetici
Из книг Гаряева П.П. можно сделать вывод, что уже давно никто из генетиков не воспринимает ТЕОРИЮ Дарвина. Из книг следует, что нужно кардинально менять весь уклад современной жизни. СМИ и ТВ нас калечат. И если раньше об этом говорили православные, то теперь с удивлением и учёные. Наука, которая говорила, что Бога нет, ЕГО же и обнаружила.
Это действительно может кого-то вдохновить на модернизм и смешение научного представления о Боге и Слове с рационалистическим.
У архимандрита Рафаила (Карелина) есть интересные замечания по поводу ограничений и подводных камней рационалистического подхода в исследовании духовного мира, в т.ч. в связи с работами А.И. Осипова, Св. Луки(Войно-Ясенецкого) и др.
Я бы обратила внимание и на А.Ф. Лосева. Правда, его метод не назовешь чисто рационалистическим.
Но есть книги наших ученых Российской Федерации: П.П.Гаряев, 2009 г., «Лингвистико-Волновой геном. Теория и практика», П.П.Гаряев «Волновой генетический код», Издательство «ИЗДАТЦЕНТР» 1997г, П.П. Гаряев «Волновой геном», Издательство «Общественная польза» 1993г. Обратите внимание на год издания книг - т.е. осознания материала.
Св. отцы говорят, что Бога можно познать умом - вот из указанных книг это и следует. Да, заглянули учёные глубже в ДНК и увидели Творца. ДНК сравнивают с текстами - со СЛОВОМ, (замысел Творца). Так прямо об этом и пишут. Правда, понимание Творца у учёных своё. Читайте, думайте, рассказывайте. Уже многое открыто.
Остался один не выясненный вопрос, камни бы возопили если бы дети Авраамовы молчали.(по памяти воспроизвожу цитату из писания). Насчет функционирования неживой материи.